Summary
The difference in the nature of codon–anticodon ambiguity seen between bacterial initiation and elongation lies not only in the position of the codon at which it occurs (5′ v 3′, respectively), but in the partial or absolute nature of the interactions (only some AUG and GUG etc. codons initiate and insert fMet, whereas all AUU and AUC codons will recognize the Phe-tRNA with anticodon, 3′-UAG-5′. It can be accounted for by the quite different molecular interactions occurring at the ribosomal P-site (for initiation) and the A-site (for elongation) and the importance of the base-pairing between the Shine & Dalgarno (RBS) region of the mRNA for initiation.
Wobble in context
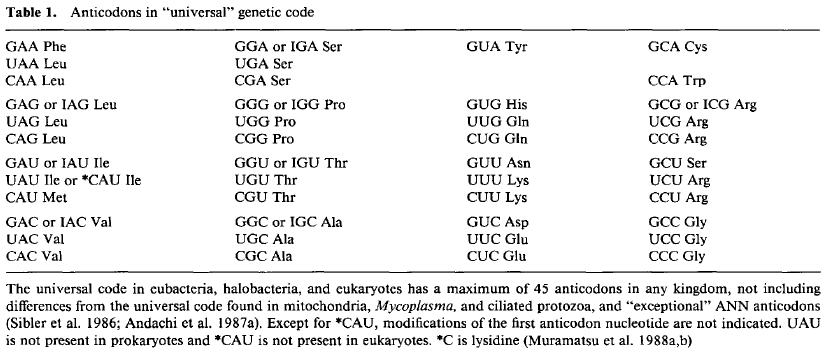
Wobble was the term applied by Crick in 1966 to the non-standard base-pairing between the 3′ position of mRNA codons and the 5′ position of anticodons of certain tRNAs. To avoid violating the genetic code Crick assumed strict base pairing at the first two codon positions and asked the question “If one sRNA [old term for tRNA] codes both XYU and XYC [because of experimental evidence that this might be the case for Phe codons], how is it done?”. And then, given the many ways bases might be paired added the constraint “How many base pairs are there in which glycosidic bonds occur in a position close to the standard one?”.
On the basis of geometry Crick predicted that anticodon 5′-U might be able to pair with a codon 3′-G (in addition to the standard A), and anticodon 5′-G with codon 3′-U (in addition to the standard C). He — unwisely in my opinion — referred to these predictions (and his prediction for inosine, I) as ‘rules’. The predictions are summarized below, together with the actual pattern that has since emerged.
It should be noted that although the two predictions involving G=U base pairs would appear reciprocal, that involving an anticodon 5′-U does not occur except in mitochondria, were it is incorrect in that the interactions are not restricted to Crick’s ‘rules’.
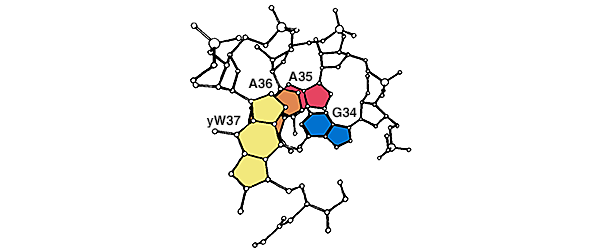
The subsequent elucidation of the structure of phe-tRNA by Quigley and Rich in 1976 suggested why the wobble occurred at the 5′ position of the anticodon. This position (G34 in my modification of their figure, below) lacks the base-stacking that would be expected to constrain the interactions at the other two positions of the anticodon, and which was aided by a modified base (yW37) 3′ to the anticodon.
As the amino acid, methionine, has only a single codon (AUG) in the genetic code, one would expect it to have a tRNA that does not allow wobble at the 5′-codon position. This turns out to be the case:
Both the tRNAmet for elongation and initiation have
the anticodon 3′-UAC-5′.
In summary: 5′-codon wobble allows mRNA to be decoded accurately by fewer than the 61 amino acid codons, and one can speculate that its emergence involved evolution of both the structures of tRNAs and the ribosomal A-site.
Initiation and decoding ambiguity
In bacteria (eubacteria) there are two main differences in codon recognition during translation from that in elongation.
- There is a single specific tRNA species involved which, uniquely, can interact with initiation factors and be bound to the P-site of the ribosome.
- Interaction is required between the 16S rRNA of the 30S ribosomal subunit and a polypurine sequence 5′ of the initiation codon (the Shine & Dalgarno sequence or ribosome-binding site) in addition to the codon–anticodon interaction.
To adopt an extreme position, the situation in initiation may almost be thought of as one in which the fMet-tRNA is bound at the P-site of the small ribosomal subunit waiting for mRNA to bind, in contrast to elongator Met-tRNA binding to an A-site with the elongation codon AUG. The energy provided by the Shine–Dalgarno interactions may thus be enough to allow initiation if one of the initiation codon bases is incorrect. This raises at least two questions:
- What advantage would allowing such ambiguous recognition have?
- Why is the ambiguity in the first codon position and how does it operate in molecular terms?
I have no answer to the first question. Perhaps it has none, but is tolerated because it has no disadvantages either. The second question is the crux of the poster’s concerns.
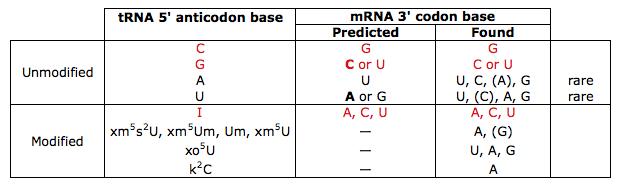
First, ambiguity at the 3′-position of the codon would not be expected on grounds of structure or precedent: the table above shows anticodon 5′-C only base-pairs with codon 3′-G.
Second, the anticodon loop of initiator tRNAmet differs from that of elongator tRNAmet in having an internal-facing rather than an external-facing disposition (Woo et al., 1980). Furthermore it lacks a modified base 3′ to the anticodon.
Third, the interactions between initiator and elongator tRNAs with the rRNA at the A- and P-sites, respectively are quite different (Berk et al. 2006), conceivably allowing more flexibility at the 3′-anticodon position.
Despite the fact that the predicted anticodon-U/codon-G wobble never occurs at the A-site, it is reasonable to suppose that structural differences allow it at the P-site, explaining the second bacterial initiation codon, GUG. As regards UUG, U=U base-pairs were considered by Crick but regarded as “rather close together”. Presumably in certain contexts this is not too energetically unfavourable to be prevented.
Finally…
The poster asked:
Am I interpreting this correctly and there's no "Crick" wobble in the AUG > GUG > UUG start codon, but rather another wobble-like mechanism that explains the start codons' first nucleotide wobble-like behavior?
To which I would reply that:
There is not and cannot be any wobble at the 3′-position of either initiation or elongation AUG codons of bacteria because it would involve non-standard base-pairing of the anticodon C. To the extent that the ambiguity at the 5′-position of the alternative initiation codons used (GUG>UUG) follows a pattern consistent with the most likely non-standard base-pairing predicted by Crick, it would seem to involve base-pairing achieved by movement (wobble) of the 3′-anticodon residue U of initiator fMet-tRNA.